





第三節(jié) 載脂蛋白基因的多態(tài)性
《動脈粥樣硬化》
在人類進化過程中,,基因的DNA序列在逐漸演變。這種演變大都發(fā)生在氨基酸編碼子以外的區(qū)域,,稱為“中性”變化,,不導致臨床疾病的發(fā)生。但是,,基因的氨基酸編碼區(qū)以及基因表達的調(diào)控區(qū)的DNA序列改變時有發(fā)生,。導致蛋白質(zhì)的合成量改變或基功能的異常。載脂蛋白結(jié)構(gòu)基因或其調(diào)控區(qū)DNA序列的突變引起致高脂血癥或早發(fā)動脈粥樣硬化已大量見諸于醫(yī)學文獻,。新的載脂蛋白基因的多態(tài)性不斷地被發(fā)現(xiàn),。
檢測載脂蛋白基因的遺傳變異通常使用從外周血白細胞中提取的DNA為起始材料,通過聚合酶鏈反應(yīng)擴增基因DNA的產(chǎn)量,,嗣后,,或采用變性梯度凝膠電泳(denaturing gradientgel electrophoresis, DGGE)顯示由基因編碼胺基酸區(qū)DAN序列突變所致的單股DNA構(gòu)象的多態(tài)性;或使用等位基因特異的寡核苷酸探針雜交法(allele-specificoligonuc leotide hybridization,ASO)探測突變的等位基因;或測定限制性片段長度多態(tài)性(restriction fragment length polymorphism, RFLP)鑒定基因DNA序列的突變;或直接對某基因的片段進行DNA序列測定,,與正常的相應(yīng)的DNA序列比較,,檢測有否突變發(fā)生。這幾種方法為較常使用的方法,。有關(guān)這些方法的詳細介紹可參見本書第十六章的生物化學檢驗技術(shù)篇,。本節(jié)主要介紹ApoB,ApoE,Apo(a),。ApoAⅠ基因的多態(tài)性,。對其他載脂蛋白基因的多態(tài)性僅以舉例的方式略述。
一、ApoB基因的遺傳變異與多態(tài)性
ApoB基因的遺傳變異與多態(tài)性分以下幾種類型進行介紹:①RFLP;②導致血漿膽固醇水平降低的基因突變;③導致血漿膽固醇升高的基因突變;④其他類型的多態(tài)性,。
1.RFLP
據(jù)報道,,載脂蛋白B基因的限制性片段長度多態(tài)性至少375種。后來發(fā)現(xiàn)有些多態(tài)性并非存在,,純屬DNA序列測定錯誤,。表4-3列舉了10種RFLP。產(chǎn)生這些RFLP的突變或發(fā)生在ApoB基因上游啟動子區(qū)域,,或發(fā)生在內(nèi)含子區(qū)域,,或在外顯子區(qū)域,。
表4-3 ApoB基因限制性片段長度多態(tài)性舉例
惹限制性內(nèi)切酶酶切位點突變等位基因出現(xiàn)頻率酶切部位存在或缺失
AvaⅡ距第1外顯子上游4Kb0.20缺失
MspⅠ啟動子區(qū)內(nèi),,-265bp0.21缺失
ApaLⅠ第4外顯子,cDNA的417bp0.36缺失
HincⅡ第4內(nèi)含子內(nèi),,距第3外顯子3'末端3334bp0.12存在
PvuⅡ第4內(nèi)含子內(nèi),,距5外顯子5'末端4523bp0.08存在
AluⅠ第14外顯子內(nèi),cDNA的1981bp0.48缺失
BalⅠ第20內(nèi)含子內(nèi),,距第21外顯子5'末端146bp0.50
XbaⅠ第26外顯子內(nèi),,cDNA的7674bp0.40-0.50存在
MspⅠ第26外顯子內(nèi),cDNA的11040bp0.12缺失
EcoRⅠ第26外顯子內(nèi),,cDNA的12670bp0.20缺失
(源自:Young S G. Circulation 82:1583,1990)
表4-3所列發(fā)生在外顯子內(nèi)的突變,,除XbaⅠ限制性片段長度多態(tài)性外,其余均導致氨基酸改變,。已有很多研究者致力于研究常見的ApoB基因PFLP與高膽醇血癥,、高甘油三酯血癥硬化的相關(guān)關(guān)系,但發(fā)現(xiàn)令人信服的相關(guān)關(guān)系的報道不是很多,,還有的結(jié)果相互矛盾,。正如Young指出的那樣,有的研究所使用的樣本量太小;有的研究對照組的選擇不合規(guī)范;有的研究基于不同的種族,,忽視了正常的種族差異,。很顯然。在以后的相關(guān)研究中,,盡可能選用大樣本量,,合理的選擇對照組,考慮不同種族的正常遺傳變異無疑對確證某些ApoE基因的RFLP與脂蛋白代謝紊亂的特殊表型以及早發(fā)動脈粥樣硬化的關(guān)聯(lián)尤為重要,。
2.導致血漿膽固醇水平降低的ApoB基因突變
經(jīng)過許多實驗室的努力,,一些ApoB基因的無義突變或移碼突變導致家族性密度脂蛋白血癥均已鑒定(見表4-4)。
表4-4 ApoB基因突變導致短ApoB的合成
ApoB的長度DNA水平上的突變短ApoB在血漿脂蛋白中的分布
B-2在第5內(nèi)含因子中,,G顛換成T無
B-9T替代C,,使翻譯在411號密碼子后終止無
B-25在第21外顯子內(nèi)有694bp缺失無
B-27.6在第24內(nèi)含子內(nèi),剪接部位突變HDL,LDL
B-29在第4152核苷酸上,,T替代C無
B-31cDNA核苷酸4480缺失HDL及密度大于1.2g/cm3部分
B-32在密碼子1450上,,T替代CHDL,LDL
B-32.5在4631核苷酸上,,G替代THDL及密度大于1.21g/cm3部分
B-37cDNA核苷酸5391至5394段缺失VLDL,,LDL,HDL
B-39cDNA核苷酸5591缺失VLDL,,LDL
B-40cDNA核苷酸5693至5694段缺失VLDL,,LDL,HDL
B-46在cDNA核苷酸6381上,,T替代CVLDL,,LDL,HDL
B-49.6在cDNA核苷酸6963上,,T替代CVLDL,,LDL,HDL
B-52.8cDNA核苷酸7295缺失VLDL,,HDL
B-54.8cDNA核甘酸7539缺失在cDNA核苷酸7655上,,T替代CVLDL,HDL
B-61在核苷酸8525處,,有37bp缺失VLDL,,HDL
B-67cDNA核苷酸9327缺失LDL,VLDL
B-74.7cDNA核苷酸10366缺失LDL,,VLDL
B-82在核苷酸11411上,,A替代CVLDL,HDL
B-86cDNA核苷酸11840缺失VLDL,,HDL
B-87cDNA核苷酸12032缺失VLDL,,HDL
B-89cDNA核苷酸12309缺失VLDL,HDL
(源自:Kane ,J.P.,and Havel, R.J.In:The metalbolic and molecular bases of inherited disease,edited by Scriver, C.R.et al, Vol Ⅱ,p1868,1995,McGraw –Hill,Inc.,New York,U.S.A)
家族性低低密度脂蛋白血癥的患者大多為雜合子,,即他們攜帶一個正常的ApoB等位基因和一個變異的ApoB等位基因,。純合子以及復合雜合子患者少見。短ApoB合成的患者血漿中ApoB的濃度較低,,僅為正常人ApoB等位基因產(chǎn)生的ApoB-100濃度的2%~10%,,其機理尚未完全明了。有人發(fā)現(xiàn),,含有ApoB87和ApoB89的LDL比含ApoB-100的LDL更易于被LDL受體攝取,。亦有人推測在雜合子患者中,由一個正常ApoB-100等位基因編碼的ApoB-100與兩個正常ApoE等位基因編碼的ApoE組成的脂蛋白顆粒比正常的脂蛋白顆粒多含有1倍的ApoE,,故前者易于被LDL攝取清除,。這些均有助于解釋一部分短ApoB合成患者體內(nèi)低濃度血漿ApoB及低濃度血漿膽固醇的機制有等進一步努力。不難想到的是,家族性低低密度脂蛋白血癥患者血漿中LDL膽固醇低,,產(chǎn)生冠心病及動脈粥樣硬化的危險性也降低,。已有一些報道證實,這些患者不產(chǎn)生動脈粥樣硬化且壽命增長,。故有人認為,,對慣常食用高飽和脂肪、高膽固醇飲食的北美人群,,攜帶合成短ApoB的突變基因非但無害,,反而有益于健康。
3.導致血漿膽固醇升高的基因突變,。
Brown和Goldstein的經(jīng)典研究,,即LDL受體基因的突變,引起LDL清除障礙,,LDL在血漿中蓄積,,導致早發(fā)動脈粥樣硬化,這是家族性高膽固醇血癥的主要病因,。這些患者的血漿總膽固醇濃度高于7.76mmol/L。然而有相當一部分人的血漿膽固醇濃度在6.47~7.76mmol/L之間,,與正常人相比為輕中度升高,。長期以來,人們一直推測ApoB基因遺傳變異可合成有缺損的ApoB蛋白質(zhì),,后者與LDL受體親和力下降,,引起LDL清除障礙,LDL在血漿中蓄積,。這種推測于1989年得到了證實,。Soria等人發(fā)現(xiàn)了家族性ApoB100缺損。這些病人在其Apob cDNA第10708個核苷酸位置上,,G轉(zhuǎn)換成了A,,導致ApoB100的等3500個氨基酸精氨酸變換成了谷氨酰胺。這一突變使其LDL與LDL受體的結(jié)合力下降,,產(chǎn)生高膽固醇血癥,。據(jù)估計,這種突變在人群中的發(fā)生率與LDL受體的突變率接近,,即1/500,。
另一家族性ApoB100缺損是由ApoB基因第10800核苷酸位置上的C轉(zhuǎn)換成T所致。這一突變ApoB內(nèi)第3531個氨基酸精氨酸變成了半胱胺酸,。8個患者的平均血漿LDL膽固醇水平在6.78mmol/L,,而8個未患ApoB基因突變的親戚為3.88mmol/L。患者LDL對LDL受體的親和力僅為正常對照組的39%,。由于C至T的轉(zhuǎn)換,,產(chǎn)生一個新的限制性內(nèi)切酶切割部位(NsiI),這將有助于在大范圍人群內(nèi)檢測這種突變發(fā)生率。
4.其他類型的多態(tài)型
距ApoB基因3'末端500個核苷酸堿基對區(qū)域,,有一DNA片段構(gòu)成可變動數(shù)目的半聯(lián)重復(variable number of tandem repeats. VNTR)已發(fā)現(xiàn)ApoB基因3'末端這種VNTR所致的多態(tài)性與心肌梗塞的發(fā)作有關(guān)聯(lián),。
ApoB信號肽由27個氨基酸組成。在高加索種族人群中發(fā)現(xiàn)了兩種多態(tài)性:一種為插入等位基因型,,編碼27個氨基酸;另一種為缺失等位基因型,,編碼24個氨基酸。Visvikis 等人發(fā)現(xiàn)缺失等位基因型與高血漿ApoB 水平,、LDL膽固醇水平,、LP(a)水平以及總膽固醇水平有關(guān)。最近Kammerer等人在34家共686名美籍墨西哥人中發(fā)現(xiàn)編碼ApoB信號肽的三種等位基因:SP-24,,SP-27,,SP-29,它們分別編碼24,,27與29肽,。攜帶SP-24等位基因的純合子其血漿ApoB與LDL膽固醇的濃度顯著高于SP-27純合子。SP-29雜合子的ApoB與LDL膽固醇水平比SP-24純合子還高,。
二,、ApoE基因的多態(tài)性
ApoE在膽固醇與甘油三酯代謝中起著重要作用。在正常人群中,,10%以上的個體間血漿膽固醇水平的著異可歸究于ApoE基因的多態(tài)性,。主要的ApoE等位基因有ApoE2、ApoE3和ApoE4,。ApoE3在人群中的出現(xiàn)率最高,,被認為較常見的ApoE基因。ApoE2與Ⅲ型高脂蛋白血癥相關(guān),,而ApoE4在老年性癡呆病(Alzheimer's disease)患者中的出現(xiàn)率較高,。這后一發(fā)現(xiàn)開拓了人們的視野,即ApoE不僅在脂類代謝中至關(guān)重要,,而且在其他的代謝過程中也扮演重要角色,。已有報道,ApoE參與組織損傷的修復,、免疫調(diào)節(jié)以及調(diào)控細胞生長與分化,。相信隨著時間推移,更多新的ApoE功能及新的ApoE基因多態(tài)性會被發(fā)現(xiàn),。
表4-5列舉了一些已鑒定的ApoE基因的遺傳變異,。表中同時列出了密碼子及其編碼的氨基酸的改變,、特定ApoE基因變異型首先被鑒定出的那個民族、出現(xiàn)頻率,、相關(guān)的高脂血癥型以及遺傳方式,。
表4-5 ApoE多態(tài)性
多態(tài)性名稱1變異密碼 子核苷酸改變氨基酸改變2種族背景分布率3高脂血癥型4遺傳方式
常見的ApoE基因型
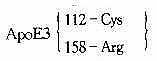
---所有民族41.9~91.1%-
ApoE4112TGC→CGCCys→Arg所有民族6.4%~36.8%N/HC
ApoE2158CGC→TGCArg→Cys大部分民族0.0%~14.5%N/FD陷性
與FD相關(guān)的多態(tài)型
ApoE0第3內(nèi)含子A →G,G缺合成兩種短肽美洲黑人14FD陷性
3592bp-60失移碼突變合成60肽德國人3FD陷性
ApoE4-費城13GAG →AAGGlu →Lys拉丁美洲人7FD陷性
145CGT →TGTArg →Cys拉丁美洲人 陷性
ApoE3-Leiden112TGC →CGCCys →Arg荷蘭人>40FD顯性
21bp重復7個氨基酸患聯(lián)重復
ApoE1127GGC→GACGly→Asp高加索人>25FD未知
158CGC→TGCArg→Cys
ApoE2-Christchurch136CGC→AGCArg→Ser未報道3FD未知
ApoE3112TGC→CGCCys→Arg拉丁美洲人6FD顯性
142CGC→TGCArg→Cys
ApoE3--kochi145CGT→TGTArg→His日本人4FD未知
ApoE2145CGT→TGTArg→His未報道3FD未知
ApoE1- Harrisburg146AAG→GAGLys→Glu日本人高加索 人8FD未知
ApoE2146AAG→GAGLys→Gln荷蘭人美國人>40FD顯性
ApoE3華盛頓210TGG→TAGTrp→終止密碼美國白人3FD陷性
ApoE2 Fukuoka224CGG→CAGArg→Gln日本人1FD未知
與非FD高脂血癥相關(guān)的多態(tài)型
ApoE53GAG→AAGGlu→Lys日本人0.5%HC
13GAG→AAGGlu→Lys法籍加拿大人6HC/HTG
ApoE2-Dunedin228CGC→TGCArg→Cys未報道2HTG
ApoE2236GTG→GAGVal→Glu荷蘭人8HTG
ApoE7-Suita244GAG→AAGGlu→Lys日本人0.5%~0.8%HC/HTG
245GAG→AAGGlu→Cys
ApoE3112TGC→CGCCys→Arg荷蘭人3HTG
251CGC→GGCArg→Gly
ApoE1158CGC→TGCArg→Cys荷蘭人2HC
252CTG→GAGLeu→Glu
與高脂血癥無關(guān)的多態(tài)型
ApoE4-+-Ferburg28CTG→CCGLeu→Pro德國高加索人0.4%~0.9%N
112TGC→CGCCys→Arg
ApoE4112TGC→CGCCys→Arg荷蘭人5N
274CGC→CACArg→His
ApoE4+296AGC→CGCSer→Arg荷蘭人5N
ApoE584CCG→CGGPro→Arg美國白人0.2%N
112TGC→CGCCys→Arg
ApoE3-Freibaug42ACA→GCAThr→Ala德國高加索人1N
ApoE399GCG→ACGAla→Thr美國人1N
152GCC→CCCAra→Pro
ApoE2134CGG→CAGArg→Gln荷蘭人5N
1.除費城(Philadelphia)與華盛頓(Washington)兩地名外,其他的地名均未翻譯過來;
2.氨基酸代號:Cys-半胱氨酸;Arg-精氨酸;Glu-谷氨酸;Lys-賴氨酸;Gly-甘氨酸;Asp-天門冬氨酸;Ser-絲氨酸;His-組氨酸;Gln-谷氨酰胺;Trp-色氨酸;Val-纈氨酸;Leu-亮氨酸;Thr-蘇氨酸;Ala-丙氨酸;Pro-脯氨酸;
3:分布率:加百分號的分布率是根據(jù)檢測1000人以上的結(jié)果計算的;未加百分號的數(shù)值代表測出的攜帶ApoE多態(tài)型(包括純合子與雜合子)的人數(shù);
4:縮寫:N(Normal,正常)HC(hypercholesterolemia,,高膽固醇血癥);FD(familialdysbetalipoproteinemia,家族性異常低密度酯蛋白血癥,,含Ⅲ型高脂蛋白血癥)。
(源自:Knijff et al. Hum. Mutation4:181,1994)
三,、Apo(a)基因的多態(tài)性
血漿Lp(a)的濃度與Apo(a)蛋白質(zhì)的大小成反比,,即Apo(a)的分子量愈大,則Lp(a)的濃度愈低;Apo(a)的分子量愈小,,則Lp(a)的濃度愈高,。Apo(a)的分子量在400~800KDa之間。按照Apo(a)在聚丙烯凝膠電脈時相對于ApoB100(分子量為520KDa)的遷移率,,Utermann等人于1987年將Apo(a)歸類為六種多態(tài)型;F比ApoB100遷移快;B與ApoB100遷移率相等;S1,,S2,S3,,S4比ApoB100遷移慢?,F(xiàn)已證實。Apo(a)蛋白質(zhì)的多態(tài)型是由Apo(a)基因中Kringle-4的數(shù)目變化而引起的,。Kring-4基因片段含有幾個稀少的限制性酶切部位(NotI,SfiI.KspI,Sval,KpnI),,這為加速篩選Apo(a)基因的多態(tài)性提供了幫助。表4-5列舉在奧地利北部Tyrolean區(qū)高加索人群中用KpnI作工具所測得的RFLP與1-4Kringe的數(shù)目,、Apo(a)蛋白質(zhì)的大小以及血漿Lp(a)濃度的關(guān)系。
表4-6 Apo(a)Kpni 片段與Apo(a)蛋白質(zhì)多態(tài)型,、Kringle-4功能區(qū)數(shù)目以及血漿Lp(a)濃度之關(guān)系
表現(xiàn)分子量(KDa)KpnI片段(Kb)Kringle-4數(shù)目等位基因 數(shù)目平均Lp(a)濃度(mg%)KpnI片段頻率(%)
F<45037-4911-131-3*<0.2
B~50055-6614-164-661.71.2
S1~55072-8217-197-934.43.8
S2~60088-9920-2210-1224.511.2
S3~650105-11623-2513-1510.213.7
S4>700121-21026-4216-32<5.770.1
*:樣本量太小,資料缺如,。(源自:Utermann,G.In:The metabolic andmolecular bases of inherited disease.7th ed, Vol.Ⅱ,eds.Scriver,C.R.etal.p1897,McGraw-Hill.Inc.,New York,1995)
由表4-6可看出,血漿Lp(a)的濃度亦與Kpn片段的大小成反比,。這為血漿Lp(a)與Apo(a)多態(tài)型的大小成反比從基因DNA水平提供了直接的證據(jù),。
Scana等人在Kringle-4第37個重復區(qū)(KIV-37,又稱為KringleⅣ-10)檢測出了兩種突變:一種突變導致第72個氨基酸被精氨酸取代;另一種突變導致第66個氨基酸蘇氨酸被蛋氨酸取代,。前者致使其Lp(a)不能與賴氨酸結(jié)合,,且血漿Lp(a)濃度降低。后者似乎與Lp(a)濃度無關(guān),。Mooser等人報道在距第1第外顯子5’端1.3Kb區(qū)或內(nèi),,由5個核苷酸[(TTT-TA)n]構(gòu)成的VNTR與其血漿Lp(a)濃度相關(guān)。此外,。亦有報道Apo(a)基因5’端側(cè)翼區(qū)DNA序列的多態(tài)性控制血漿Lp(a)的濃度,。
四,、ApoAⅠ基因的多態(tài)性
ApoAⅠ為HDL的主要結(jié)構(gòu)蛋白。ApoAI基因突變可導致異常的ApoAⅠ蛋白質(zhì)的合成,。有些異常的ApoAⅠ影響HDL的代謝,。已報道的阻礙ApoAⅠ合成的基因突變有重排、缺失,、無義突變等方式,。這些突變的純合子患者表現(xiàn)出極低HDL膽固醇水平和早發(fā)冠心病。目前已發(fā)現(xiàn)至少有20種不同的ApoAI結(jié)構(gòu)基因的點突變導致氨基酸轉(zhuǎn)換(見表4-7),。其中兩種突變(精氨酸173→半胱氨酸和脯氨酸165→精氨酸)與低HDL膽固醇水平相關(guān),,但并不引致早發(fā)冠心病。
Talmud等人發(fā)現(xiàn)ApoAⅠ基因啟動子區(qū)域內(nèi)距轉(zhuǎn)錄起始部位第75個堿基(-75bp)A置換G的突變與高ApoAI及HDL水平相關(guān),。這種突變后的等位基因稱為A等位基因,,而含G的稱為G等位基因。Saha等報道A等位基因在新加坡華人中出現(xiàn)的頻率(0.27)高于高加索人(0.12),。Wang等人發(fā)現(xiàn),,ApoAⅠ基因第1內(nèi)含子5'末端,第83堿基C轉(zhuǎn)換成T或第84堿基G轉(zhuǎn)換成A,,致使MspI酶切點喪失,,這種突變與高HDL膽固醇相關(guān),比上述的-75堿基A置換G突變與高HDL膽固醇的相關(guān)還強,。
表4-7 人體ApoAI基因點突變導致氨基酸改變
氨基酸改變1密碼子2核苷酸點突變
Pro3→ArgCCCC→G
Pro3→HisCCCC→A
Pro4→ArgCCCC→G
Arg10→LeuCGAG→T
Gly26→ArgCGGCG→C
Asp89→GluGATCT→G,T→A
Asp103→AsnCGACG→A
Lys107→0
Lys107→MetAAGA→T
Glu136→LysCGAGG→A
Glu139→GlyGAGA→G
Pro143→ArgCCAC→G
Glu147→ValGAGA→T
Ala158→GluGCGC→G
Pro165→ArgCCCC→G
Glu169→GlnCGAGG→C
Arg173→CysGCGCC→T
Arg177→HisCGCG→A
Glu198→LysCGAGG→A
Asp213→GlyGACA→G
1:氨基酸代號可參見表4-5腳注;第1排氨基酸右上角的數(shù)值表示其在ApoAⅠ一級結(jié)構(gòu)上的位置;2:加底線的核苷酸為點突變部位,。(源自:Eckardstein et al.J.Biol Chem.265:8612,1990)
五、其他載脂蛋白基因的多態(tài)性
與以上介紹的ApoB,、ApoE,、Apo(a)以及ApoAI的基因一樣,其他載脂蛋白的基因也存在多態(tài)性,,以下簡要地列舉一些例子,。
1.ApoAⅠ-CⅢ-AⅣ基因限制性片段長度多態(tài)性
在ApoAⅠ-CⅢ-AⅣ基因叢內(nèi),使用XmnⅠ,、ApaⅠ,、MspⅠ、PstⅠ,、SstⅠ,、XbaⅠ、TaqⅠ和PvuⅡ限制性內(nèi)切酶已檢測出10多種RFLP,。盡管這些多態(tài)性大多由編碼氨基酸以外的DNA序列突變所致,其中有些RFLP的攜帶者與增加冠心病的危險相關(guān),。比如用限制性內(nèi)切酶PstⅠ消化從人群中搜集的DNA后。用ApoAⅠcDNA探針印跡DNA可測得3.3Kb的DNA片段(稱為P2等位基因)與2.2Kb的DNA片段(稱為P1等位基因),。P1等位基因在正常人群中出現(xiàn)率較高,。P2等位基因在低HDL膽固醇患者中的出現(xiàn)率高于對照組的6倍,。來自英國與美國的報道亦證實,雜合子P1P2攜帶者血漿ApoAⅠ與HDL膽固醇的水平顯著地低于P1P2基因型攜帶者,。這提示在ApoAⅠ-CⅢ-AⅣ基因叢中,,PstⅠ酶切點的多態(tài)性,P2等位基因,,與降低血漿ApoAⅠ和HDL的水平以及誘發(fā)冠心病有關(guān),。最近,Dallinga-Thie等人在美國脂類研究雜志上報道,,用限制性內(nèi)切酶XmnⅠ與MspⅠ在ApoAⅠ-CⅢ-AⅣ基因叢內(nèi)檢測出的稀有等位基因與高血漿膽固醇,、甘油三酯、ApoB以及LDL膽固醇水平相連,。這些結(jié)果顯示,,ApoAⅠ-CⅢ-AⅣ基因叢參與調(diào)節(jié)血漿膽固醇及甘油三酯的代謝。
2.ApoEⅠ-CⅠ-CⅡ基因限制性片段長度多態(tài)性
位于第19對染色體上的ApoEⅠ-CⅠ-CⅡ基因叢也有多態(tài)性的存在,。檢測出其RFLP的限制性內(nèi)切酶包括:HpaⅠ,、TaqⅠ、BglⅡ,、DraⅠ,、NcoⅠ以及BglⅡ。Klasen等人報道,,HpaⅠ限制性片段長度的多態(tài)性與易發(fā)Ⅲ型高脂血癥有關(guān),。然而HpaⅠ限制性片段長度的多態(tài)性與E2/E2基因型無關(guān)接關(guān)聯(lián)。這說明HpaⅠ的RFLP是不同于ApoE基因的一種獨立的遺傳因子,,其突變點尚未闡明,。
3.ApoAⅡ基因的突變
Deeb等報道了在一對日本姐妹中檢測出的家族性ApoAⅡ缺陷癥。冠狀動脈造影顯示這對姐妹未患血管疾病,。盡管免疫生化檢驗測不出血漿ApoAⅡ水平,,患者HDL膽固醇在正常范圍。DNA分析顯示,,患者ApoAⅡ基因內(nèi)第3內(nèi)含子剪接供體部位的第一個堿基G突變成為A,從而阻礙了內(nèi)含子從初級轉(zhuǎn)錄物中剪除掉的過程,。這無疑是導致ApoAⅡ缺陷的原因,。
4.ApoAⅣ基因的多態(tài)性
常見的ApoAⅣ等位基因編碼ApoAⅣ的第360與347個氨基酸,分別為谷氨酰胺和蘇氨酸,。當ApoAⅣ基因的第360密碼子上第3個堿基T置換G后,,正常的第360氨基酸位置上的谷氨酰胺被代之為組氨酸,當?shù)?47密碼子上第1個堿基T置換A后,,蘇氨酸347→絲氨酸的變異即發(fā)生,。關(guān)于這兩種ApoAⅣ基因多態(tài)性對脂類代謝的影響尚無一致的結(jié)論,。另一種突變,即密碼子127的突變可致HincⅡ酶切位點的喪失,。最近Kamboh等人對西伯利亞中部的馴鹿牧民檢測中發(fā)現(xiàn),,沒有HincⅡ酶切位點等位基因的攜帶者血漿甘油三酯水平高于HincⅡ酶切位點等位基因的攜帶者。
5.ApoCⅡ基因的多態(tài)性
載脂蛋白CⅡ基因的突變有四種類型被鑒定:a.無義突變;b.起動密碼子的堿基突變;c.第2內(nèi)含子剪切供體部位突變;d.由堿基插入或缺失所致的移碼突變,。這些突變患者的血漿ApoCⅡ水平顯著降低或缺失,。其遺傳方式為常染色體隱性遺傳。
6.ApoCⅢ基因的多態(tài)性
已發(fā)現(xiàn)有三種ApoCⅢ基因多態(tài)性與高甘油三酯血癥有關(guān):a.3175核苷酸C顛換成G;b.3206核苷酸T顛換成G;c.3206G等位基因,。Li等人不久前在臨床研究雜志上報道,,ApoCⅢ基因啟動子內(nèi)-482與-455堿基的突變與發(fā)生高甘油三酯血癥有關(guān)。作者推測這種突變正發(fā)生在受胰島素抑制DNA序列范圍內(nèi),。突變后的ApoCⅢ基因啟動子擺脫了胰島素的抑制調(diào)控,,ApoCⅢ的轉(zhuǎn)錄大大加強,ApoCⅢ的合成增加,。這可能是人群中發(fā)生高甘油三酯的一個主要因素,。
7.ApoD基因的多態(tài)性
Vijayaraghavan等人檢測了57個肥胖病人57名對照者ApoD基因TaqⅠ酶切位點限制性片段長度多態(tài)性:2.2與2.7Kb等位基因。他們發(fā)現(xiàn)2.2Kb等位基因在肥胖者中的分布顯著地高于正常者,。這包括含有2.2Kb等位基因的雜合子與純合子,,提示顯性遺傳方式。作者認為,,ApoD基因TaqⅠ限制性片段長度多態(tài)性可作為研究肥胖病的一個遺傳標志,。
8.ApoH基因的多態(tài)性
ApoH基因第247密碼子突變,致使第247個氨基酸纈氨酸被亮氨酸取代,。因為此突變使RasⅠ酶切位點喪失,,故這種限制性片段長度多態(tài)性很易檢測。最近Kamboh等調(diào)查西伯利亞中部馴鹿牧民中三種ApoH等位基因ApoH1,、ApoH2,、與ApoH3的分布與血漿脂質(zhì)水平的關(guān)系,他們發(fā)現(xiàn)ApoH等位基因與男子低甘油三酯水平相關(guān),。而與女子高甘油三酯水平相關(guān),。而女子高甘油三酯水平相關(guān)。ApoH基因的多態(tài)性與脂質(zhì)代謝的關(guān)系還有待于更廣泛地,、更深入地探討,。
9.ApoJ基因的多態(tài)性
Kamboh暨同事報道,在對美國白種人,、黑人,、美洲印第安人、愛斯基摩人,、新幾內(nèi)亞人及尼日利亞人6個民族共985名受試者的檢測中,。他們發(fā)現(xiàn)了ApoJ有三種多態(tài)型,,即ApoJ1、ApoJ2和ApoJ3,。ApoJ1存在于所有受試者基因內(nèi),。ApoJ2僅存于美國黑人與尼日利亞人黑人,出現(xiàn)頻率分別為24%與28%,。ApoJ3僅從一美國黑人受試者測出,。造成此多態(tài)性的突變尚未見報道。這三種“等位基因”似乎對總膽固醇,、LDL膽固醇,、HDL膽固醇、VLDL膽固醇,、甘油三酯的水平?jīng)]有顯著的影響,。
結(jié)論
隨著分子生物學技術(shù)應(yīng)用于脂類代謝與動脈粥樣硬化的研究領(lǐng)域,幾乎所有被命名的載脂蛋白的cDNA與基因被分離與鑒定,。有關(guān)載脂蛋白基因多態(tài)性的報道與日俱增,。人們對脂蛋白代謝紊亂從分子缺陷的水平進行認識取得了長足的進展。然而,,各種載脂蛋白的生理功用尚未完全闡明,。近年來發(fā)現(xiàn)ApoE4與老年性癡呆有關(guān),揭示了ApoE的新的功用就佐證了這一點,。對各種載脂蛋白的基因的各種多態(tài)性在所有種族間的分布進行系統(tǒng)的檢測與分析還有待于進一步努力,。一些新的、簡易的,、準確的檢測方法也有待于開發(fā),。這些研究可望為載脂蛋白與動脈粥樣硬化發(fā)病機制的探討以及對動脈粥樣硬化與冠心病的防治尋求新的措施開拓新的前景。
(葉永清)
- 標簽:
- 高脂血癥